
Genomics/Epigenomics
Category: Abstract Submission
2: Genomics/Epigenomics II

Jim N. Jarvis, MD
Professor of Pediatrics
Jacobs School of Medicine and Biomedical Sciences at the University at Buffalo
Buffalo, New York, United States
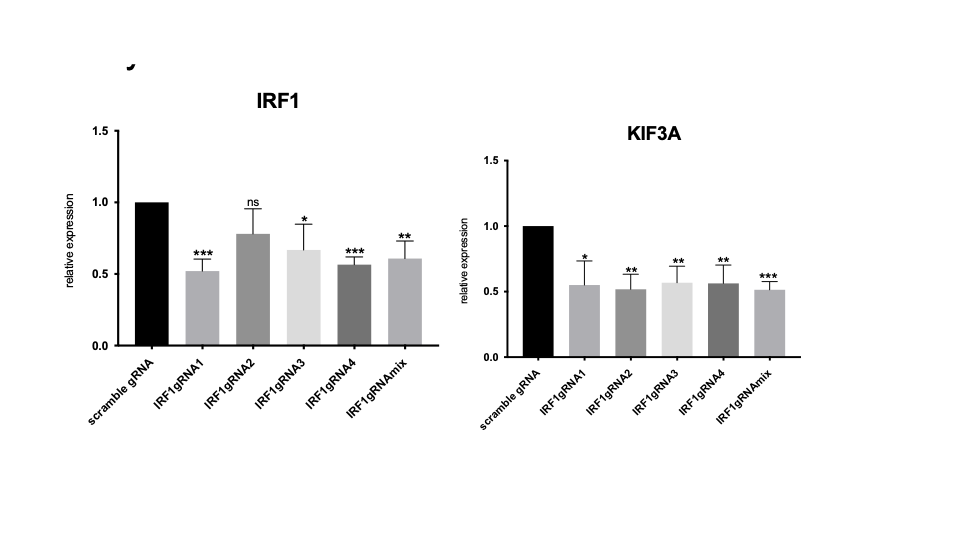 The bar graph showing results obtained by ablating the intergenic enhancer harboring rs2706385 and rs2706386 on the IRF1 locus using CRISPRi with dCas-KRAB and 4 gRNAs tiled across the enhancer region or used together (“gRNAmix”). Ablating this enhancer reduces expression of both IRF1and KIF3A. *p < 0.05; **p < 0.01; ***p < 0.001.Ns=Not significant. Attenuating this enhancer had no effect on the expression of GDF9 (not shown), another gene within the same TAD as this enhancer.
The bar graph showing results obtained by ablating the intergenic enhancer harboring rs2706385 and rs2706386 on the IRF1 locus using CRISPRi with dCas-KRAB and 4 gRNAs tiled across the enhancer region or used together (“gRNAmix”). Ablating this enhancer reduces expression of both IRF1and KIF3A. *p < 0.05; **p < 0.01; ***p < 0.001.Ns=Not significant. Attenuating this enhancer had no effect on the expression of GDF9 (not shown), another gene within the same TAD as this enhancer.